Наши предки по телу — кто они и сколько их было: некоторые проблемы научно-богословского синтеза
Опубликовано: 10 сентября 2024
Автор

Павлов Алексей Михайлович
Кандидат биологических наук
Пролог
Существуют области знания, которые являются зоной внимания как ученых, так и богословов. Когда-то таким общим полем было строение Солнечной системы. Галилей был сторонником гелиоцентризма и имел немало неприятностей от того, что в современное ему католическое богословие было включено положение о центральном положении нашей планеты во вселенной. Только в ноябре 1979 г. папа Иоанн Павел II официально признал, что инквизиция в 1633 г. совершила ошибку, вынудив ученого отречься от его научных взглядов.
История с отношением к геоцентризму показала, что создание богословских моделей, которые, описывая мир после грехопадения, противоречат достоверно установленным научным фактам и концепциям, приводит в конечном счете к краху таких моделей. При этом важно отметить, что, если исходить лишь из анализа текста Библии, были очень веские основания принять геоцентрическую модель (по тексту светила «обращаются» — «восходят», «шествуют по небосводу», «заходят», «останавливаются» во время битвы при Гаваоне). И только наблюдения, возможность которых обусловило развивающееся естествознание, помогли понять, что это — лишь удобные для восприятия образы. Поэтому утверждение, что для понимания Библии достаточно лишь самого ее текста, по крайней мере в том, что касается описания падшего мира, не всегда верно. Так или иначе, понимание слов Библии невозможно без наблюдения тех объектов или процессов, которые обозначены словами в Священном Писании. И в некоторых случаях наука может дать достоверное знание об этих объектах и процессах.
В случае с вопросом о строении Солнечной системы можно относительно легко включить данные естественно-научных наблюдений в христианскую мировоззренческую картину. Однако при описании истории человечества вопрос о соотнесении богословского и научного знания не получается разрешить так же легко.
Введение
Между описаниями истории человечества, которые дают наука и Священное Писание, есть, на первый взгляд, существенные противоречия:
1) Библия повествует об актах творения двух бессмертных совершенных людей, их грехопадении и, вследствие этого, — появлении смерти. Современное человечество — потомки этой пары.
2) Наука говорит об эволюционном совершенствовании множества изначально смертных гоминид, которые постепенно обретали те свойства, которые присущи современному человеку.
Существует несколько мировоззренческих моделей, включающих в картину мира и библейскую историю, и эволюционные взгляды на происхождение человека, наиболее известные представлены в таблице 1. Каждая из моделей включает определенное описание человеческой истории на нашей планете, которая может быть сопоставлена с современными научными данными. В нашей работе будут анализироваться те особенности истории человечества, которые определяют применимость той или иной «модели соотнесения».
|
Модель соотнесения |
Как описывается история человечества |
|
Креационизм молодой Земли С. Ю. Вертьянов[1] |
История человечества начинается с божественного акта сотворения двух совершенных людей — Адама и Евы, изгнанных из Рая, который находился на той же Земле, где и сейчас обитает человечество (т. е. тело современного человека не имеет эволюционной связи с приматами). Исторический период существования человечества на планете — около 6000 лет. При этом тело человека существенно не меняется. |
|
Теистический эволюционизм А. В. Кураев[2] А. В. Гоманьков[3] А. В. Григорян[4] |
Современные люди по телу — потомки древних приматов. Постепенное совершенствование тела Homo происходило в течение миллионов лет. Образ Божий появляется у Homosapiens в результате божественного воздействия. В разных вариантах теистического эволюционизма в этот момент людей могло быть двое или много. |
|
Альтеризм Еп. Василий (Родзянко)[5] Н. С. Серебряков[6] Мумриков О., свящ.[7] В. В. Иваненков[8] А. В. Храмов[9] |
Для альтеризма характерно резкое разграничение состояний мира до и после грехопадения. Первозданный мир был совершенен, «хорош весьма», после грехопадения он резко изменился (появились смерть, страдания, энтропия). В концепции еп. Василия история падшего мира начинается с Большого взрыва, современное человечество — «разлетевшиеся осколки Адама». Во взглядах других авторов история человечества на нашей планете начинается с Адама и Евы, изгнанных из Рая, который находится после грехопадения в иной области бытия и недоступен для научного исследования, причем реальная возможность появления человечества всего от двух людей не обсуждается с научной точки зрения. В варианте концепции, развитой о. Олегом Мумриковым, для жизни на нашей планете при изгнании из Рая людям даны «кожаные ризы» — их тела претерпели изменения, в результате божественного предведения актуализировались детали, создающие видимость происхождения Homo эволюционным путем, сделав человека одним из многих биологических видов планеты. По мнению В. В. Иваненкова, первые люди после грехопадения попадают из Рая в мир «проклятой Земли», обладая телами, ставшими тленными в результате грехопадения. |
Табл. 1. Наиболее известные модели соотнесения Библейской истории и научных данных (указаны только некоторые авторы, выражающие ту или иную «модель соотнесения»)
Надо отметить, что признание эволюции тела человека входит в часть «моделей соотнесения». Поэтому необходимо сформулировать конкретные вопросы об особенностях истории человечества, принципиально важных для верификации этих или других возможных «моделей соотнесения»:
1) Является ли современный человек по телу эволюционным потомком приматов?
2) Возможно ли, что современное человечество происходит от только двух прародителей?
3) Какие качества отделяют «настоящего человека» от животных?
4) Появились ли качества
«настоящего человека» у Homosapiens или у более ранних видов?
В настоящей работе будут обсуждаться только первые два вопроса. В процессе рассмотрения этих вопросов будут обсуждаться критерии достоверности, но об одном из них стоит сказать сразу. Если мы видим на полу отпечатки, идентичные по контуру и всем деталям подошвам ботинок, то делаем вывод, что эти отпечатки оставили ботинки. Здесь критерий достоверности таков: если мы видим многочисленные факты, которые можно объяснить как следствие определенных процессов, причем случайное их появление невероятно, то мы считаем, что эти процессы действительно происходили. Кстати, этот критерий использует и судебная практика, принимая как доказательство отпечатки пальцев или структуру ДНК.
Также стоит отметить, что ответы
на поставленные выше вопросы не будут однозначно свидетельствовать о
правильности какой-то из упомянутых «моделей соотнесения», так как, в принципе,
возможны и иные богословские решения. Но если те принципиальные особенности
истории человечества после грехопадения, которые включены в модель, не
соответствуют реалиям нашего мира, то такая модель явно неадекватна. Также
автор не считает правильным априорное пренебрежение мнениями святых отцов
Церкви, которым могло быть открыто то, что рассудку ученого, наблюдающего лишь
современный материальный мир, никогда не откроется.
Является ли современный человек по
телу эволюционным потомком
древних приматов?
Возможная в наше время детальная расшифровка ДНК делает возможным сравнение геномов с целью сравнительного и исторического анализа. Среднее расхождение последовательностей генома человека и высших приматов (шимпанзе и горилла), оцененное по 53 фрагментам ДНК, составляло 1,24–1,62%[10]. В теле человека и современных приматов не обнаруживается никаких качественных различий их природы на материальном уровне (сходство строения, состава белков, хромосомного набора). Однако это сходство можно, в принципе, интерпретировать как сходство по причине «одного Творца», так что сам по себе этот аргумент недостаточен.
Разные модели «соотнесения» в описании истории человека различаются, в частности, в том отношении, что в известных вариантах альтеризма и креационизма молодой Земли люди на нашей планете появляются в какой-то момент времени и не имеют эволюционной связи с приматами. В модели теистической эволюции такая связь предполагается. Сравнить адекватность этих моделей возможно, сравнивая «эволюционные деревья» приматов, полученные разными способами.
«Эволюционное дерево» можно построить по сходству состава геномов. «Эволюционные деревья», построенные по ископаемым останкам гоминид и на основе сходства геномов, совпадают. Но все же остается возможность, что это положение человека на «эволюционном дереве» объясняется не следствием эволюции, а сходством его генома (который мог твориться Богом независимо) с геномом высших обезьян.
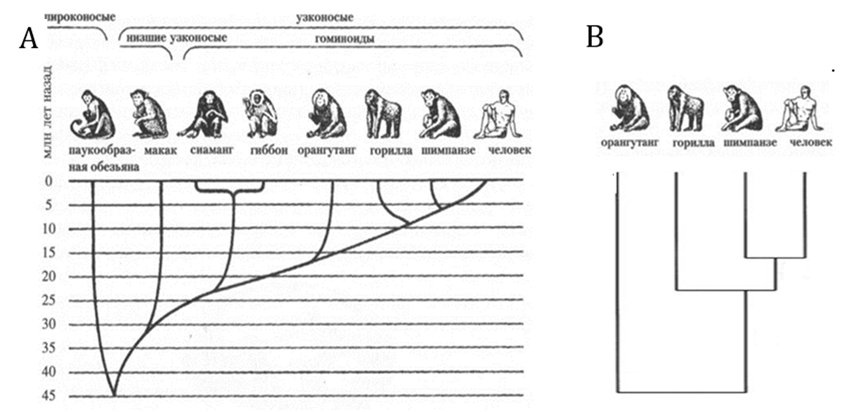
Рис. 1. А. Эволюционная ветвь высших приматов (Anthropoidea) по данным о ископаемых останках. Показаны лишь линии, представители которых сохранились поныне[11]. В. «Эволюционное дерево» высших приматов, построенное на основе сходства ДНК[12]
Более весомым доказательством было
бы обнаружение генетических маркеров, сравнивая наличие которых у разных видов,
можно было бы восстановить ход эволюции. Такими маркерами могут считаться
вставки в ДНК, которые передаются в ряду поколений. На эту роль подходят, в
частности, так называемые эндогенные ретровирусы (ERV). В науке доминирует точка
зрения, что эта вставка в ДНК — копия РНК ретровируса, образовавшаяся в
результате воздействия так называемых ретровирусов[13].
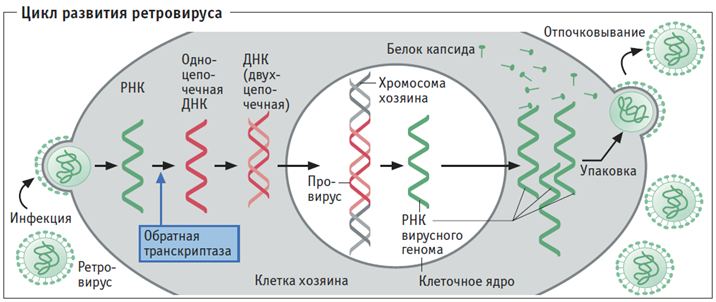
Рис. 2. Цикл развития ретровируса
Ретровирус — это покрытая белковой капсулой цепочка рибонуклеиновой кислоты (РНК), на которой записана необходимая для его воспроизводства генетическая информация. Ретровирус проникает в клетку, затем на его РНК синтезируется молекула ДНК с аналогичной информацией и встраивается в молекулу ДНК атакованной клетки. Эта вставка называется «эндогенный ретровирус» (ERV); эндогенные ретровирусы, свойственные Homo, называют HERV. Затем обычным для клетки путем, с использованием информации «вирусной вставки» в ДНК, синтезируются необходимые для вируса белки и РНК. Но клетки могут перевести «вирусную вставку» (ее называют провирусом) в своей ДНК в неактивную форму. Если эта вирусная ДНК попала в половые клетки, она будет передаваться в ряду поколений.
Анализируя количество общих для разных видов ретровирусных вставок у ныне живущих видов, можно, таким образом, понять, как выглядело их эволюционное древо. Ведь однажды встроенная в ДНК копия ретровируса никуда из этого места генома обычно не перемещается. Учитывая размер геномов позвоночных (>109 пар нуклеотидов) и относительно равномерный и случайный характер внедрений ретровирусов, вставка одинаковых последовательностей ERV в одном и том же месте крайне маловероятна[14]. Существует расчет, сравнивающий вероятности случайного внедрения ретровирусов в ДНК одного примата с вероятностью гипотезы, что они унаследованы этим приматом от общих предков[15]. При этом учитывается, в частности, недавно открытое явление, что вероятность внедрения ретровирусов в некоторые точки ДНК может быть несколько выше, чем в другие. По результатам расчетов, гипотеза случайного внедрения менее вероятна на 46 порядков величин (единица с 46-ю нулями). Надо отметить, что в расчетах везде в случае неопределенности принимались оценки величин, минимизирующих вероятность «гипотезы общего предка». Поэтому, если мы видим у двух видов идентичные ретровирусные последовательности ERV1 и ERV2, но не видим их у третьего, то первые два вида (A и B), скорее всего, ближе друг к другу (Рис. 3).
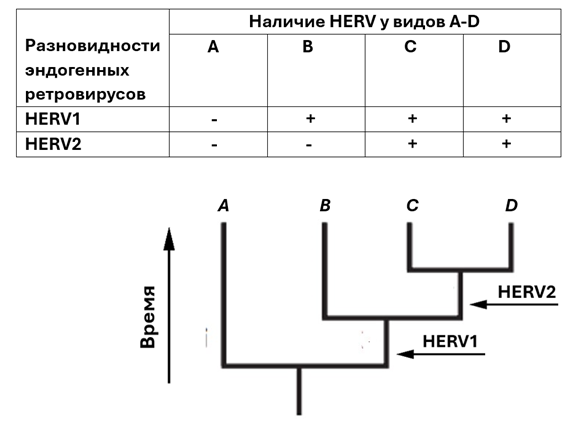
Рис. 3. Принцип построения
эволюционного древа на основе выявления ретровирусных последовательностей (HERV) в геноме животных[16]. Стрелками показано
время внедрения разновидностей ретровирусов
Однако, поскольку некоторые эндогенные ретровирусы могут иногда обладать специфической генетической активностью, в том числе и полезной для организма — носителя этих вставок[17], сторонники появления человеческого тела (такого же, как и тело современных людей в том мире, в котором мы сейчас обитаем) в результате специального божественного творческого акта считают, что вставки в геном, которые интерпретируются как «эндогенные ретровирусы», сотворены в числе прочих генов. В таком случае можно было бы ожидать, что эти вставки состоят исключительно из функционально полезных участков. Однако надо отметить, что у большинства эндогенных ретровирусов, свойственных человеку, активна лишь часть участков, а остальные — блокированы[18]. Современный взгляд на эволюционную динамику HERV, основанный на множестве проведенных исследований, обобщенных в работе А. А. Буздина с соавторами[19], состоит в том, что на ранних стадиях после внедрения клетки-хозяева воспринимают HERV как чужеродный генетический элемент и, вероятно, подавляют его путем целенаправленного метилирования и мутаций. Однако на более поздних стадиях, когда уже накоплено значительное количество мутаций и когда ретровирусные гены нарушены, регуляторный потенциал HERV может быть высвобожден. При этом эндогенные ретровирусы в определенных ситуациях вызывают или усугубляют патологии: рак, воспаления, инфекционные заболевания и др[20]. Странно предполагать, что всемогущий Бог вставил в геном человека для благих целей что-то очень похожее на вирусы, но с блокированными вирусными генами, при этом в основном бесполезное, иногда — частично полезное, а иногда — вредное. Причем вставил именно так, что эти вставки соответствуют эволюционной картине, аналогичной эволюционным картинам, полученным другими способами.
Построение «эволюционного древа» на основе сравнения набора ретровирусных вставок или их фрагментов у разных видов — не единственный способ построить «эволюционное древо» на основе анализа ДНК. Существует также подход, основанный на анализе отличий в так называемых «длинных концевых повторах» (LTR) у разных видов приматов и человека (Рис. 4А). Дело в том, что, когда ДНК одной из групп ретровирусов внедряется в ДНК клетки животного, по краям этой вставки появляется несколько дополнительных нуклеотидов, которые и образуют LTR, находящиеся по обе стороны вирусной ДНК (их называют 3’LTRи 5’LTR). Причем когда вирус внедряется в ДНК хозяина, в связи с особенностями механизма транскрипции в составе 3’LTR и 5’LTR образуются идентичные по составу блоки. В них могут с течением времени происходить мутации — изменения в последовательности «букв генетического кода» — нуклеотидов. На основе сходства и различия мутаций LTR у разных видов приматов (причем отдельно — для 3’LTR и 5’LTR) можно построить картину эволюции этих видов. Достаточно очевидно, что точечные мутации в LTR не могли бы образовывать закономерной картины эволюционных изменений, вдобавок сходной для 5’LTR и 3’LTR последовательностей каждого эндогенного ретровируса в случае отсутствия эволюционного преемства рассматриваемых видов.
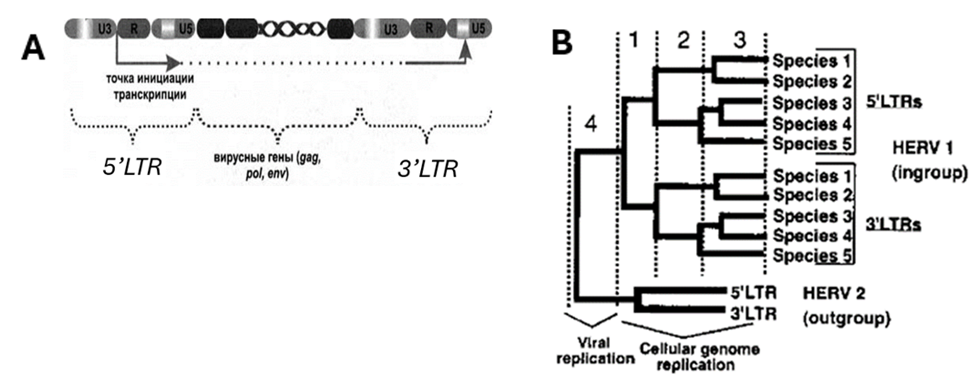
Рис. 4. Анализ мутаций в «длинных концевых повторах» (LTR) эндогенных ретровирусов как инструмент построения «эволюционного древа». А — пример строения «эндогенного ретровируса» с двумя «длинными концевыми последовательностями» 5’LTR и 3’LTR[21]. В — пример гипотетического эволюционного древа, восстановленного с помощью анализа генетического состава «длинных концевых повторов» (5’LTR и 3’LTR) двух гипотетических эндогенных ретровирусов (НERV-1 и НERV-2). Эволюционная история, о которой свидетельствует накопление мутаций в 5’LTR и 3’LTR, начинается после внедрения вируса в геном животного (фаза 1). Поскольку мутации происходят в каждом из «длинных концевых повторов» — 5’LTR и 3’LTR — независимо, можно выстраивать эволюционные деревья на основе данных по каждому из LTR, проверяя таким образом надежность реконструкции[22]
И наконец,
третий источник информации о филогении группы — носителя ретровирусов — это
количество мутаций, накопленных в LTR определенного вида, которое может быть
своего рода «эволюционными часами» — мерой времени, прошедшего с момента
внедрения вируса в геном[23].
Дело в том, что из-за механизма обратной транскрипции два LTR, скорее всего,
будут идентичны в момент объединения ДНК-генома с образованием провируса.
Поэтому сравнение последовательностей двух LTR на концах генома эндогенных
ретровирусов позволяет оценить их относительный возраст. Чем старше провирус,
тем больше различий, вероятно, будет присутствовать. А если известна частота
мутаций, то можно оценить и абсолютный возраст.
Рассмотрим те исследования, в которых какими-то из перечисленных выше трех способов получали сведения о филогении приматов и человека на основе исследования ретровирусных вставок. В работе Барбулеску с соавтарами[24] эволюционное древо человека и приматов в явном виде не представлено, но там приведены данные о нахождении десяти ретровирусных последовательностей в их геномах (Табл. 1). Это позволяет построить эволюционное древо для упомянутых видов (Рис. 1). Оно вполне соответствует представлениям о филогении приматов и человека, полученных на основании анализа костных остатков, обнаруженных в земных слоях, приведенных в монографии Л. Б. Вишняцкого[25] (Рис. 5).
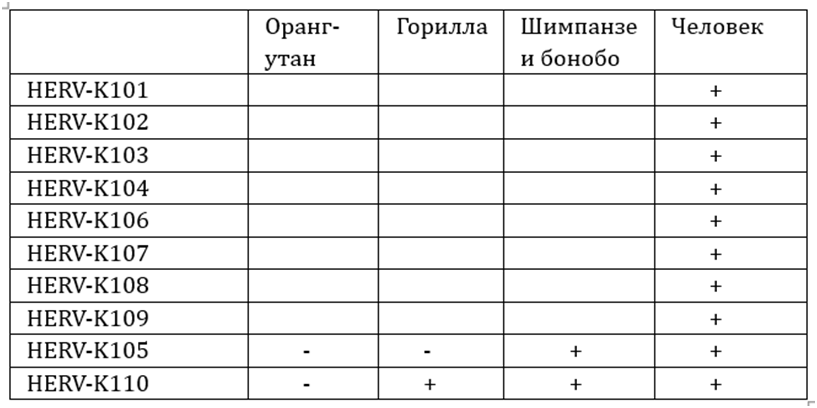
Табл. 1. Наличие 10 разновидностей
эндогенных ретровирусов у приматов и человека по данным Barbulescuetal., 1999[26]
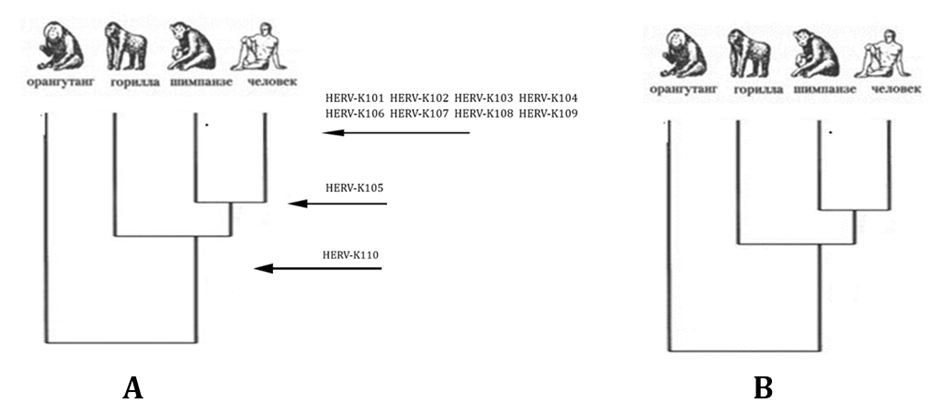
Рис. 5.
Эволюционные древа приматов и человека, построенные по данным Barbulescuetal., 1999 о наличии эндогенных ретровирусов HERB-K (A) и на основании анализа костных остатков[27]
(В)
Надо отметить, что в работе Барбулеску также приведены данные о количестве различий в «длинных концевых повторах» (LTR) ретровирусов, позволяющие оценить их относительный возраст. Различия в LTR вирусов, свойственных только человеку, находились у семи разновидностей в пределах от нуля до шести, у одной — составляли 17. Два провируса, общих для человека и современных человекообразных обезьян (HERV-K105 и HERV-K110), имели наибольшее количество различий (36 и 38), что согласуется с представлениями об их значительном возрасте по положению на эволюционном древе.
Масштабное исследование, основанное на анализе девятнадцати разновидностей длинных концевых повторов (LTR) у приматов и человека, было проведено Ю. Б. Лебедевым с сотрудниками[28]. По принципу наличия и отсутствия разновидностей LTR было реконструировано эволюционное древо (Рис. 6А). Оно идентично древу, построенному на основе костных останков (Рис. 6В). Также была оценена скорости мутирования LTR-последовательностей и, исходя из данных об их изменчивости, рассчитано время внедрения каждой из разновидностей ретровирусов в геном приматов. Для 19 групп LTR возраст появления исходного варианта ретровирусной вставки составил от 3,5 до 53 млн лет. В большинстве случаев также было отмечено совпадение времени внедрения исходных ретровирусов с представлениями о филогении приматов, полученными на основе анализа LTR-последовательностей.
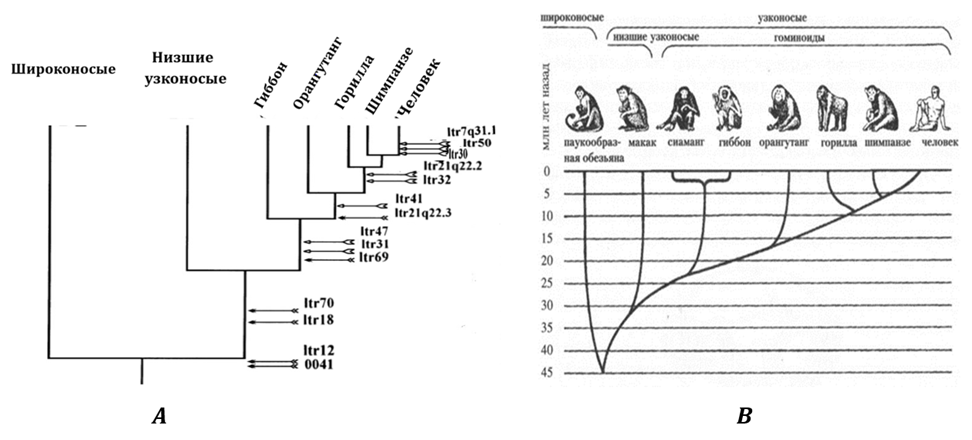
Рис. 6А. «Эволюционное дерево» высших приматов, построенное на основе: А
— анализа вставок эндогенного ретровируса человека (HERV-K) в идентичных
локусах хромосом человека и приматов[29]. В — по данным о ископаемых останках[30]
В работе Джонсона и Коффина[31] длинные концевые последовательности (LTR) семи эндогенных ретровирусов (HERV) были изученыу девяти видов приматов и у человека (при этом 5’LTR и 3’LTR анализировались отдельно). По результатам анализа нуклеотидного состава LTR были построены «деревья максимальной экономии»(MP) (Рис. 7).
Несмотря на то что последовательность нуклеотидов в LTR может и изменяться в результате различных процессов, происходящих в геноме (вставки и утраты участков ДНК, кроссинговер и др.), LTR человекa, шимпанзе и «карликового шимпанзе» бонобо, как правило, оказывались наиболее сходными по составу; LTR-последовательности гориллы и орангутанга — несколько более далекими;а африканской зеленой обезьяны (AGM) и макаки,относящихся к низшим узконосым, — наиболее далекими, что соответствует данным палеонтологии (Рис. 8).
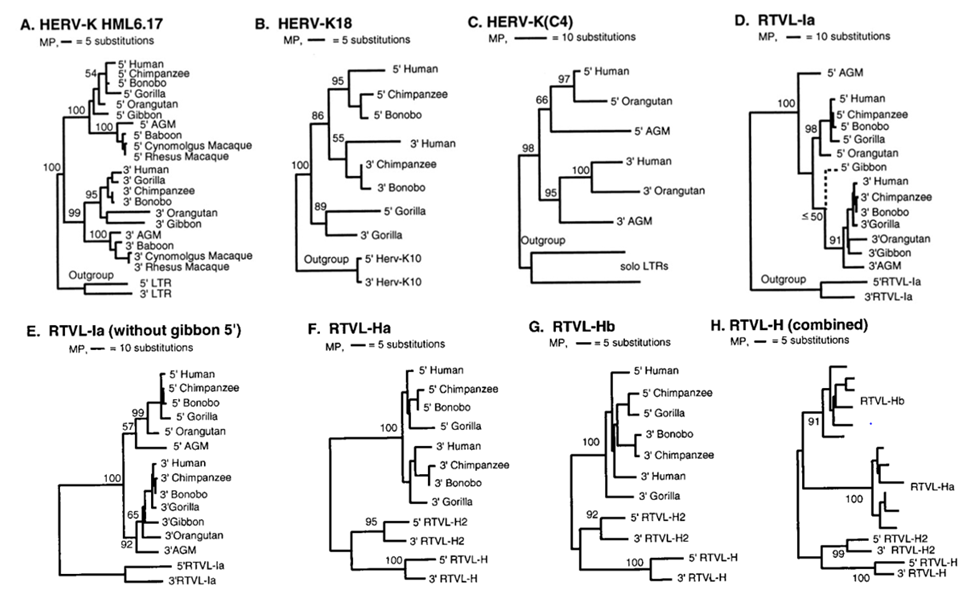
Рис. 7. Филогении семи эндогенных ретровирусов HERV на основе исследований их 5’LTR и 3’LTR-последовательностей у 10
видов приматов[32]
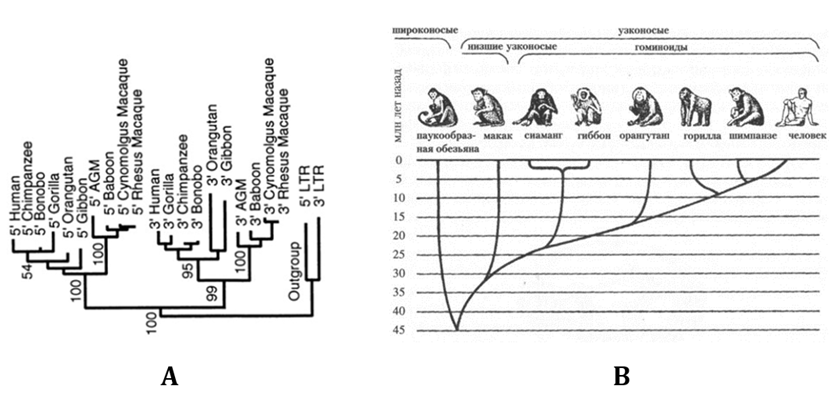
Рис. 8. «Эволюционные деревья» приматов и Homo, построенные на основе: А — данных о составе концевых
последовательностей (LTR) эндогенного ретровируса HERV-KHML6.17. (5’ и 3’ — «деревья», построенные на основе анализа 5’
и 3’ концевых последовательностей (LTR)
ретровируса)[33].
B — по данным об ископаемых останках[34]
При этом, как отмечают Джонсон и Коффин, LTR-последовательности тех ретровирусов, которые внедрились у основания «эволюционного древа», имели бóльшую изменчивость, чем у тех, которые внедрились ближе к концам «ветвей», что свидетельствует об их бóльшем эволюционном возрасте.
Итак, «эволюционные деревья»,
построенные на основании пяти групп независимых данных: четырех подходах к
анализу генома и по результатам изучения датированных костных останков приматов
и человека в основном совпадают. Могут ли такие совпадения — и картины
эволюции приматов по костным останкам, и по общему сходству ДНК, и по
результатам анализа следов ретровирусов в ДНК в трех независимых аспектах —
быть «просто случайностью»? Так как это практически невероятно, то нужно
признать, что тело человека произошло от общего с современными приматами предка
в течение непрерывной эволюционной истории.
Могло ли современное человечество произойти от двух предков,
обитавших когда-то на нашей планете
В православной информационной среде можно встретить сообщения о том, что «ученые доказали», что на Земле в определенное время жили «первые люди»: «Y-хромосомный Адам» и «митохондриальная Ева», от которых «произошло все человечество». Что же это за понятия? Действительно они идентичны «предкам всего человечества»?
Y-хромосома — это хромосома, которая у человека есть только у мужчин. Так как Y‑хромосома передается только от отца к сыну, то все современные Y‑хромосомы происходят от данного мужчины, которого называют «Y‑хромосомным Адамом». Это наиболее близкий по времени общий предок всех ныне живущих людей по мужской линии. Но и до него, и в его время жили другие люди, гены Y-хромосомы которых не дошли до потомства (Рис. 9).
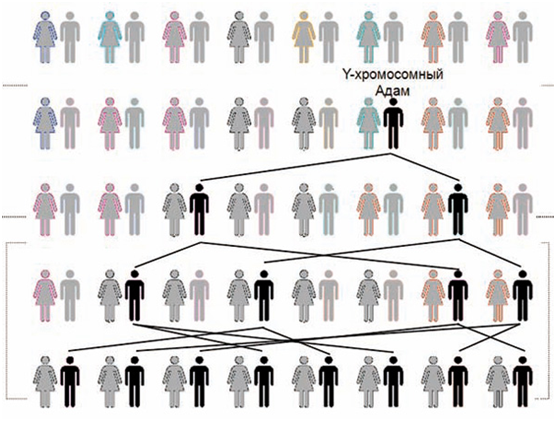
Рис. 9. «Y-хромосомный Адам» — общий предок,
опр��деленный по признаку сходства генетического состава Y-хромосомы. При наличии в каждом
поколении многих людей, через несколько поколений остаются потомки только
одного человека, которого и называют Y-хромосомным Адамом. И до него, и
в его время жили и другие люди
Митохондрии есть в каждой клетке. У каждой из них есть собственная ДНК (не та, что в ядре клетки) — мтДНК. Ребенок наследует только митохондрии матери, которые находятся в яйцеклетке. Следовательно, у каждого ребенка мтДНК — только от матери. Через отцов мтДНК не передаются. Для того, чтобы женщина передала свою митохондриальную ДНК другим поколениям, ей нужно родить дочь, а той родить ещё дочь, и так далее. Общий предок, определенный по мтДНК, — «митохондриальная Ева». Схема демонстрирует (Рис. 10), каким образом в результате генетического дрейфа разнообразие мтДНК в популяции в конце концов приходит к одному-единственному варианту, носителем которого становятся все женщины. Рождение детей разного пола и в разном количестве при передаче потомкам только через дочерей неизбежно приводит к тому, что только один вариант мтДНК остается в популяции. При этом вычисленная «митохондриальная Ева» не является единственной женщиной и не является первой в цепи предков.
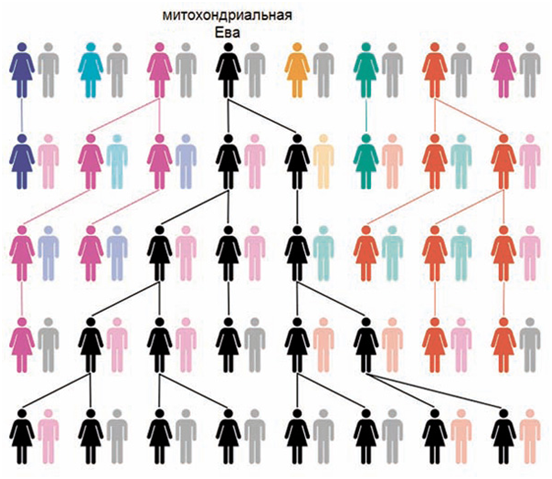
Рис. 10. «Митохондриальная Ева» — общий предок,
определенный по признаку сходства генетического состава Y-хромосомы. Показано, что, при наличии в каждом поколении многих
людей, через несколько поколений остаются потомки только одной женщины, которую
и называют «митохондриальной Евой»
Чтобы лучше понять этот процесс, его можно рассмотреть на модельном примере, предложенным А. Марковым: «Допустим, есть поколение 0, и в нем 20 женщин, каждая из которых имеет свой вариант мтДНК. В следующем поколении каждая из этих женщин производит на свет некое количество (например, от 0 до 4) дочерей (сыновьями в данном случае можно пренебречь). Из общего числа дочерей в поколении остаются снова 20. И каким-то женщинам поколения 0 не повезло: они дочерей не родили или их дочери не выжили. Таким образом, в поколении 1 разнообразие мтДНК несколько уменьшилось. Так происходит от поколения к поколению: постепенно в популяции остается все меньше вариаций мтДНК. И хотя частота каждого из 20 гаплотипов колеблется по закону блужданий, примерно к 30-му поколению из исходных вариантов остается один. В этот момент женщина, которая внесла в поколение 0 этот вариант мтДНК, становится митохондриальной Евой всей живущей популяции. Это стохастический, основанный на случайностях процесс, но исход его неизбежен: все живущие женщины популяции будут прямыми потомками по женской линии одной-единственной женщины из поколения 0»[35]. Поэтому вычисления времени существования «митохондриальной Евы» и «Y-хромосомного Адама» (которые, кстати, могут иметь разный эволюционный возраст) вообще не имеет отношения к решению вопроса о том, сколько было наших предков и когда они жили.
Возможно ли в принципе появление современного человечества от одной пары? Одним из способов оценить возможность этого является исследование влияния близкородственных скрещиваний, которые в таком случае неизбежны. Так как в предыдущем разделе было доказано, что современное человечество по телу происходит от древних приматов, нужно понимать, что при этом неизбежен «генетический груз» вредных и смертельных рецессивных мутаций, находящихся только в одной из парных хромосом. В таком случае мутация не проявляется. Но если участки ДНК, несущие этот поврежденный ген, будут представлены в обеих хромосомах, то организм ждет летальный исход или пониженная жизнеспособность
Известный исторический пример, подтверждающий опасность инбридинга у людей, — испанская династия Габсбургов. Последний из ее представителей, живший в XVII в. Карл II был потомком трех родственных пар «дядя — племянница» и смог дожить лишь до 38 лет. Карл II был не только уродлив, но и страдал от многочисленных болезней, включая дефицит гормонов гипофиза, тяжелое заболевание почек и глубокое слабоумие. До восьми лет юный король не мог самостоятельно передвигаться, а до десяти — отличить А от Б. Также он был неспособен оставить потомство, отчего династия Габсбургов и прервалась[36]. Понадобилось всего три поколения не самых близкородственных браков.

Рис. 11. Король Карл II Габсбургской династии
В работе Перес-Перейры[42] с помощью математического моделирования оценивался минимальный размер жизнеспособной популяции (МРП) животных в долгосрочной перспективе на фоне действия различных факторов, которые могут этот МРП ограничивать. Учитывалось влияние частоты мутаций, доли вредных и летальных мутаций, эффекта близкородственного скрещивания (инбридинг), естественного отбора (причем было учтено усиление естественного отбора при инбридинге). Результаты моделирования показали, что МРП близок к 500 особям для видов с умеренно высокими темпами размножения (около шести потомков в год) и 1000 особей — при более низком уровне воспроизводства. При этом на фоне отрицательного действия каких-то экологических факторов величина МРП может быть и больше.
Эмпирические оценки размеров минимально жизнеспособной популяции крупных приматов, сделанные на основе выживаемости популяций на островах разного размера, составляют несколько тысяч для Pongo (орангутан) или десятков тысяч Symphalangus (сиаманг)[43]. Франклин в обзоре литературы утверждает, что минимальный размер жизнеспособной популяции для крупных млекопитающих составляет по крайней мере 50 особей[44].
Таким образом, результаты моделирования и эмпирических научных исследований показывают, что минимальный размер жизнеспособной популяции (МРП) крупных млекопитающих, включая гоминид, по самым минимальным оценкам должен насчитывать десятки особей, хотя нередко оценки МРП и гораздо выше.
В последние годы разработаны и усовершенствованы математические методы, позволяющие оценивать численность человеческих популяций в древности на основе анализа генома. В частности, был разработан метод, при применении которого результаты расчетов не зависят от демографической модели, по которой изменялась численность популяции. Такой математический анализ, сделанный на основании частот аллелей в десяти современных африканских популяциях человека, показал[45], что в самые критические периоды падения численности (800–900 тыс. лет назад) количество наших предков по телу составляло около 1280 особей, причем нижний предел численности популяции в этот период — 730 особей (Рис. 12). Численность Homo до и после прохождения этого «горлышка бутылки» была, по расчетам, выше примерно в 75 раз. Как отмечают авторы, вычисленный ими временной интервал «горлышка бутылки» соответствует периоду малой численности имеющихся ископаемых остатков гоминин в Африке и Евразии 950–650 тыс. лет назад. При этом относительно высокая численность Homo, по данным расчетов, имела место по крайней мере 0,9–1,4 млн лет назад, когда существовал только заметно отличающиеся от Homosapiens вид — Нomoerectus[46].
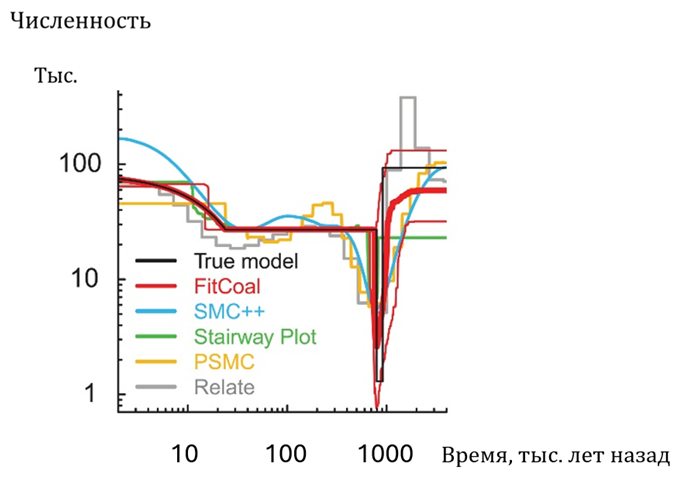
Рис.
12. Расчет динамики численности Нomo по данным о составе генома десяти современных африканским
популяций Homosapiens на основе нескольких
математических моделей[47]
Итак, с учетом рисков неблагоприятных последствий близкородственного скрещивания численность популяций Homo в любой момент истории при естественных процессах не могла быть менее нескольких десятков индивидуумов, причем данные генетического анализа свидетельствуют, что количество тех Ноmo, от которых происходят африканские популяции Homosapiens, никогда не была меньше нескольких сотен в последние 1,4 млн лет. В те времена Homosapiens не существовал, так что «креационизм молодой Земли» явно не соответствует этим научным данным.
Все варианты концепции альтеризма подразумевают, что человечество произошло от двух индивидуумов. С христианской точки зрения, невозможно представить, что появившиеся на планете люди могли бы вступать в брак с похожими на них по телу животными. Следовательно, если принять происхождение земного человечества от двух людей, то, в рамках альтеризма, нужно признать, что человечество обитает на нашей планете более чем 1,4 млн лет. В то время вид Homosapiens еще не существовал, и довольно странным выглядит обретение в качестве «кожаных риз» для совершеннейших, хотя и павших Адама и Евы тела с объемом черепа примерно в два раза меньшим, чем у современного человека.
Вопрос рассматривается серьезный, и полезным представляется определить время появления «настоящих людей» на нашей планете не только на основе данных генетики, но и по другим независимым данным. Эту возможность может предоставить анализ материальной культуры Homo, если бы удалось найти такие проявления этой материальной культуры, которые свидетельствовали бы о том, что они принадлежат «настоящему человеку» с образом Божьим.
Выводы
Научные данные по вопросу происхождения тела современного человека от древних приматов включают целый ряд согласующихся друг с другом независимых исследований, как генетических, так и палеоантропологических, выводы которых соответствуют предположению об эволюционном происхождении тела человека. Случайное совпадение этих результатов невероятно. Объяснить это можно либо эволюционным происхождением тела человека, либо тем, что Бог «имитировал» эволюционную историю нашего тела в «кожаных ризах», которые дал Адаму и Еве. В таком случае, учитывая данные многих генетических исследований, нужно признать, что Бог ввел в состав генома человека участки, идентичные фрагментам ретровирусов, которые нейтральны или даже вредны для человека.
С чисто научной точки зрения, с учетом рисков неблагоприятных
последствий близкородственного скрещивания, численность популяций Homo
в любой момент истории не могла быть менее нескольких десятков индивидуумов. При
принятии концепции альтеризма необходимо предположить, что Бог в течение многих
поколений редактировал геном человека. Данные генетического анализа
свидетельствуют, что количество тех Ноmo,
от которых происходят современные африканские популяции Homosapiens, никогда не была меньше нескольких сотен в последние 1,4 млн лет.
Тогда Нomosapiens не было, а
существовали лишь Нomoerectusc объемом мозга в два
раза меньшим, чем у современного человека. В рамках альтеристских концепций
тогда следует признать, что «кожаные ризы», данные Богом первым людям, — это Нomoerectus или даже более ранние виды.
Благодарности
Автор благодарит за обсуждение этой работы прот. Олега Мумрикова,
диак. Николая Серебрякова, канд. биол. наук В. В. Иваненкова, канд. биол. наук А.
В. Храмова, О. Соловьева.
Библиография
Василий (Родзянко), еп. Теория распада Вселенной и вера Отцов. Каппадокийское богословие — ключ к апологетике нашего времени. Апологетика XXI века. М.: Паломник, 2003.
Вертьянов С. Ю. Общая биология: Учебник для 10–11 классов общеобразовательных учреждений. М.: Свято-Троицкая Сергиева Лавра, 2012.
Вишняцкий Л. Б. Введение в преисторию. Проблемы антропогенеза и становления культуры. Курс лекций. Кишинев: Изд-во Высшей Антропологической школы, 2005.
Гоманьков А. В. Библия и природа: Эволюция, креационизм и христианское вероучение. М.: ГЕОС, 2014.
Григорян А. В. Соотнесение научной картины мира и библейской истории грехопадения. 2021. Ч. 2.
Иваненков В. В. Неадекватность моделей теистической эволюции и младоземельного креационизма: в чем причины? Часть 4. Альтернативные модели. URL: https://bogoslov.ru/article/4873408 (дата обращения: 09.12.2024).
Кураев А. В. Может ли православный быть эволюционистом. Клин: Христианская жизнь, 2006. (Андрей Вячеславович Кураев включен Министерством юстиции РФ в реестр иностранных агентов.)
Лебедев Ю. Б. Эндогенные ретровирусы человека: структурно-эволюционный анализ. М., 2004. (автореф. дис. доктора биол. наук).
Марков А. Адам, Ева и математика // Популярная механика. 2013. № 5.
Марков А. Эволюция человека. В 2 кн. Кн. 1: Обезьяны, кости и гены. М.: CORPUS, 2011.
Мумриков О., свящ. «И сделал Господь Бог Адаму и жене его одежды кожаные, и одел их»: опыт библейско-богословского осмысления реалий современной эволюционной антропологии. URL: http://www.bogoslov.ru/text/2458777.html (дата обращения: 09.12.2024).
Серебряков Н. С. Проблема соотнесения библейского повествования о творении мира и человека с научным естествознанием // «Вся премудростию сотворил еси...». М.: Труды семинара «Наука и Вера». М.: Издательство ПСТГУ. 2011. Вып. 1. С. 88–111.
Супотницкий М. В. Эволюционная патология. М.: «Вузовская книга», 2009.
Франклин Я. Р. Эволюционные изменения в небольших популяциях. Биология охраны природы. М.: Мир, 1983. C. 160–176.
Храмов А. В. Обезьяна и Адам. Может ли христианин быть эволюционистом? М.: Никея, 2018.
Alvarez G., Ceballos F. C. e. a. The Role of Inbreeding in the Extinction of a European Royal Dynasty // PloS one. N 4 (4). P. 1–7. https://doi.org/10.1371/journal.pone.0005174.
Barbulescu M. e. a. Many human endogenous retrovirus K (HERV-K) proviruses are unique to humans // Current biology. N 9 (16). P. 861–868. https://doi.org/10.1016/s0960-9822(99)80390-x.
Bond S. Endogenous retroviral insertions as evidence for evolutionary relationships among primates. URL: https://biology.stackexchange.com/questions/69076/endogenous-retroviral-insertions-as-evidence-for-evolutionary-relationships-amon (дата обращения: 09.12.2024).
Buzdin A. A., Prassolov V., Garazha A. V. Friends-Enemies: Endogenous Retroviruses Are Major Transcriptional Regulators of Human DNA // Frontiers in chemistry. T. 5 (35). P. 35–43. https://doi.org/10.3389/fchem.2017.00035.
Caballero А., Bravo I., Wang J. Inbreeding load and purging: implications for the short-term survival and the conservation management of small populations // Heredity. 2017. № 118. P. 177–185. https://doi.org/10.1038/hdy.2016.80.
Chen F., Li W. Genomic Divergences between Humans and Other Hominoids and the Effective Population Size of the Common Ancestor of Humans and Chimpanzees // American journal of human genetics. 2001. Т. 68 (2). P. 444–456. https://doi.org/10.1086/318206.
Eyre-Walker A., Keightley P. D. High genomic deleterious mutation rates in hominids. Nature. 1999. № 397. P. 344–347. https://doi.org/10.1038/16915.
Fay J. C., Wyckoff G. J., Wu C. I. Positive and negative selection on the human genome // Genetics. № 158 (3). P. 1227–1234. https://doi.org/10.1093/genetics/158.3.1227.
Harcourt A. Empirical estimates of minimum viable population sizes for primates: Tens to tens of thousands? // Animal Conservation. 2002. № 5 (3). P. 237–244.doi:10.1017/S1367943002002287.
Hu W., Hao Z., Du P. e. a. Genomic inference of a severe human bottleneck during the Early to Middle Pleistocene transition // Science (New York, N.Y.). 2023. № 381 (6661). P. 979–984. https://doi.org/10.1126/science.abq7487.
Johnson W. E., Coffin J. M. Constructing primate phylogenies from ancient retrovirus sequences // Proceedings of the National Academy of Sciences of the United States of America. 1999. № 96 (18). P. 10254–10260. https://doi.org/10.1073/pnas.96.18.10254.
Lebedev Y. B., Belonovitch O. S., Zybrova e. a. Differences in HERV-K LTR insertions in orthologous loci of humans and great apes // Gene. № 247. P. 265–277. https://doi.org/10.1016/s0378-1119(00)00062-7.
Lohmueller K., Indap A., Schmidt S. e. a. Proportionally more deleterious genetic variation in European than in African populations // Nature. 2008. № 451. P. 994–997. https://doi.org/10.1038/nature06611.
Morton N. E., Crow J. F., Muller H. J. An estimate of the mutations damage in man from data on consanguineous marriages // Proceedings of the National Academy of Sciences of the United States of America. 1956. № 42 (11). P. 855–863. https://doi.org/10.1073/pnas.42.11.855.
Pérez-Pereira N., Wang J., Quesada H. e. a. Prediction of the minimum effective size of a population viable in the long term // Research Square. 2022. https://doi.org/10.21203/rs.3.rs-1608971/v1.
Stein R., DePaola R. V. A Human endogenous retroviruses: our genomic fossils and companions // Fisiological genomics. 2023. № 55 (6). P. 249–258.
Sunyaev S., Ramensky V., Koch I. e. a. Prediction of deleterious human alleles // Human molecular genetics. 2001. № 10 (6). P. 591–597. https://doi.org/10.1093/hmg/10.6.591.
Ward E. S. W. Distribution of targets for avian retrovirus DNA integration in vivo // Genes and Development. 1994. № 8 (12). P. 1473-1487. https://doi.org/10.1101/gad.8.12.1473.